
![[Translate to Italiano:]](/fileadmin/Der_SAC/Unserer_Partner/CSS_cyan_rgb.svg)


Une plante mal connue: l'Edelweiss
Contribution à la connaissance de la végétation alpine. ( Avec une planche photographique, une carte et trois vignettes dans le texte. ) Au nombre des végétaux comptant parmi les plus représentatifs de la flore alpine, 1'«Edelweiss » peut être cité comme étant tout à la fois le plus symbolique et le moins bien connu des alpinistes non initiés aux mystères de la botanique systématique ou même de la géobotanique.
Pour appuyer ce dire, qu' il suffise de citer quelques-uns des noms populaires sous lesquels cette plante singulière est connue en diverses langues européennes; par exemple, en français: « Etoile des Alpes », « Belle étoile », « Reine des Glaciers », etc., sans compter les noms issus de préjugés erronés tels, que 1'«Etoile du Mont-Blanc » ou la « Fleur du Righi », octroyés par des tenanciers de bazar qui n' ont eu cure de s' informer si ce végétal existe bien réellement au Righi ou au Mont-Blanc, deux montagnes célèbres qui en sont totalement dépourvues. En italien, notre plante est le plus souvent désignée sous les noms de « Stella d' Italia » ou de « Fiori di roccia »; en allemand, les noms d'«Edelweiss » et d'«Alpenkönigin » sont les plus généralement usités, le premier d' entre eux surtout, qui était déjà connu dans les Alpes de Salzburg en 1784; toutefois d' autres noms moins poétiques apparaissent aussi dans la littérature bien avant cette date: c' est ainsi que dans son remarquable « Pflanzenleben der Alpen », 2eédition, publiée à Zurich en 1924, notre savant collègue de la section Uto du C.A.S., M. le professeur C. Schröter, signale, à la page 498 de cet ouvrage, le nom de « Wullblumen » attribué à l' Edelweiss dès 1561 par Aretius et 1574 par Simler; utilisée en herboristerie pour combattre les douleurs intestinales, cette herbe est alors désignée sous le nom de « Bauchwehblume », tandis qu' en remontant jusqu' au XIe siècle, elle est connue comme « Lewenwurz ». Les Anglais en font leur « Tyrolese » ou « Swiss cudweed », parfois aussi « Lion' s foot »; pour les Roumains, c' est l'«Albumela alpina », les Bulgares, « Etoile des Balkans », et les Slovènes, la « Rose du Triglav ».
En voilà assez pour l' Europe; mais si l'on songe que notre continent ne représente qu' une très petite éclaboussure de l' aire générale de l' Edelweiss, dont le foyer de dispersion occupe les vastes régions de l' Asie centrale comprises entre l' Himalaya, le Thibet et surtout la Chine nord-occidentale, avec irradiations jusqu' au Turkestan et à l' Altaï vers l' occident, la Sibérie baïcalienne et le bassin de l' Amur au septentrion, le Japon à l' orient et Formose au midi ( voir la carte de distribution ), on peut se représenter l' immensité du champ de travail que doit encore offrir ce domaine aux doctes chercheurs d' étymologie ou de folklore; nous nous bornerons à retenir le nom d'«Herbe à la diarrhée », communément appliqué à notre Edelweiss par les Sibériens, qui paraissent avoir des raisons pour apprécier davantage cette fleur sous forme de tisane qu' à titre d' appareil vexillaire juché sur un chapeau!
Enfin, pour clore cette digression nomenclaturale, il convient de rappeler que la dénomination scientifique internationale du groupe est le nom de « Leontopodium », soit « Léontopode » ou « Patte-de-lion » en français.
Tandis qu' en Europe le genre Leontopodium n' est représenté que par une seule espèce qui doit porter le nom de Leontopodium alpinum ou « Léontopode des Alpes»1 ), le continent asiatique héberge plus d' une trentaine d' espèces fort variées qui, pour la plupart, affectent un port très différent de celui de notre plante d' Europe 2 ); nous en faisons figurer quelques exemples dans notre planche photographique.
C' est par erreur ou par défaut d' observation que quelques auteurs ont attribué à l'«Edelweiss » une aire américaine et une aire australienne: les Leontopodium gnaphaloides Hieronymus ( 1900 ) et linearofolium Benth. et Hooken ( 1873 ), de l' Amérique du Sud, appartiennent sans conteste au genre Gnaphalium; le Leontopodium Meredithae F. v. M., de Tasmanie, constitue avec deux autres espèces australiennes un genre autonome, celui des Ewartia; les deux « Edelweiss » de la Nouvelle-Zélande sont dans le même cas et forment le genre Leucogenes ( L. Leontopodium et L. grandiceps ). Enfin, une plante désignée sous le nom d'«Edelweiss d' Australie », VActinotus leucocephalus Hooker, n' appartient pas même à la famille des Composées: elle se rattache aux Ombellifères tout comme notre soi-disant « Chardon bleu»Pour d' autres détails, voir le Bulletin de la Société botanique de. Genève, vol. I ( 1909 ), p. 369, et vol. II ( 1910 ), p. 236.
Cela dit, qu' il nous soit permis maintenant de passer en revue quelques détails de structure florale de notre Leontopodium alpinum, en nous aidant pour cela des figures dessinées tout exprès à l' usage de notre revue.
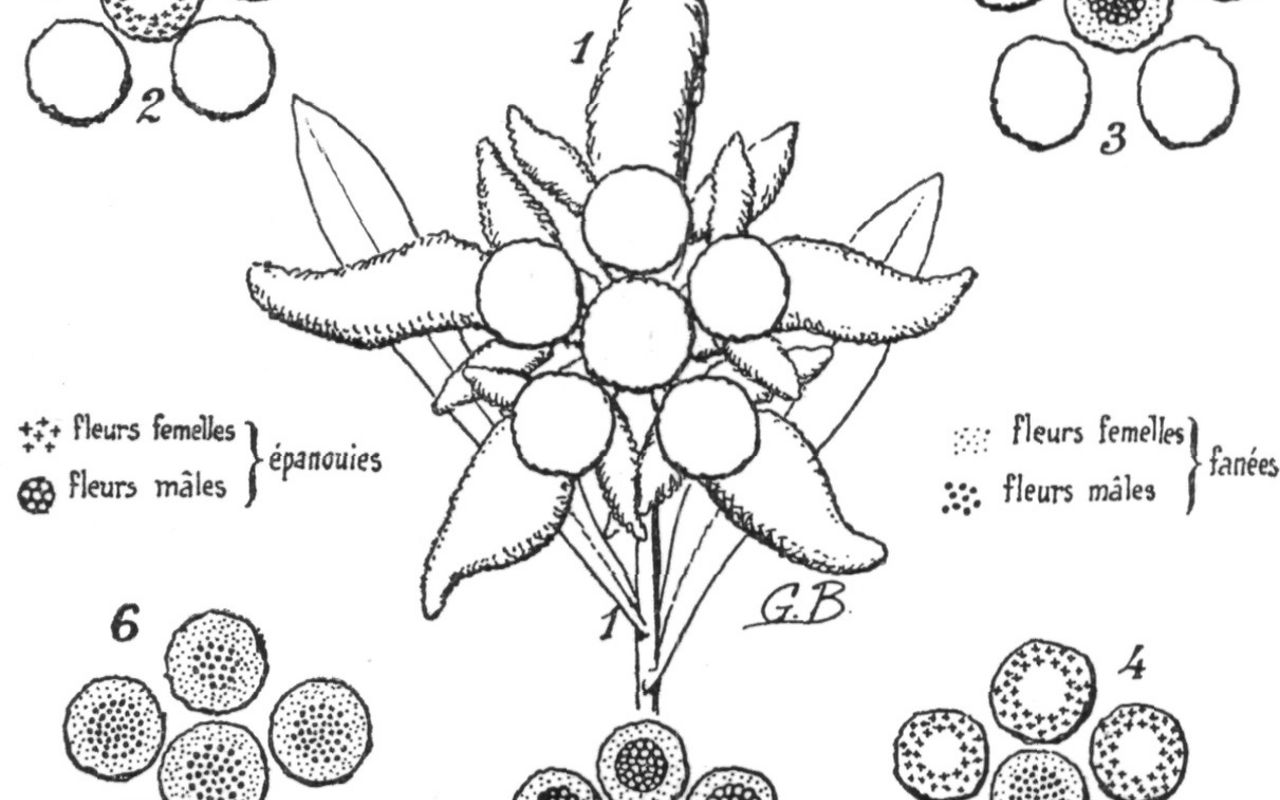
Tout d' abord, observons que les véritables fleurs de léontopode ne sont nullement les grandes étoiles feutrées de blanc et dont les rayons laineux sont fréquemment qualifiés de « pétales » par la majorité des profanes: il faut les reconnaître dans ces minuscules petits points verdâtres qui sont groupés en boules au centre de l' étoile ( fig. II: 2, 3, 4, 5 et 6 ). Ces boules, qui sont les « inflorescences en capitule » des descripteurs, constituent l' un des caractères constants permettant de reconnaître les divers membres de la grande famille naturelle des Composées; les rayons de UNE PLANTE MAL CONNUE: L' EDELWEISS.
Fig. I. Organes floraux du genre Leontopodium.
Ì: coupe longitudinale d' une tige fleurie ne présentant que des rameaux florifères simples, c'est-à-dire terminés par un seul capitule r '. 2: id. à rameaux florifères subramifiés, donnant naissance au bas du pédoncule de r' à deux ramuscules terminés par un capitule r ". 3: diagramme d' une inflorescence simple, ne présentant que des rameaux latéraux monocéphales r '. 4: id. d' une inflorescence subramifiée, à rameaux r' et ramuscules r "; dans ces quatre exemples, le capitule central r est considéré comme rameau primaire; la feuille radiale primaire est marquée al », les feuilles radiales secondaires ab ' », et les bractéoles de 3e ordre « b "; f = feuilles caulinaires. 5: feuilles du capitule, soit a écaille du péricline » glabres à l' intérieur int ( grossi X 12 ). 6: id ., laineuses sur le dos ( ext. X 12 ). 7: fleur femelle complète ( grossi X 16 ). 8: soie de l' aigrette femelle, finement scabre ( X 16 ). 9: sommet de la corolle, développé ( x X32 ). 10: fleur femelle dépourvue de son aigrette, avec ovaire glabre en a ( x 16 ). 11: id. dépourvue de sa corolle; ovaire papilleux en a ': nectaire en n; « regme » en r; épiregme en e; stigmate en st. ( X 16 ). 12: schéma d' une fleur hermaphrodite, c'est-à-dire présentant des organes des deux sexes ( cas très exceptionnel chez les Leontoimdium ); en a, ovaire fertile; n, r et e, comme chez les fleurs femelles; corolle du type mâle, à 5 divisions régulières; en i., étamines mal formées, non soudées entre elles en manchon ( x 16 ). 13: fleur male, à corolle masquée par l' aigrette ( x 16 ). 14: id ., soie de l' aigrette mâle, sensiblement épaissie au sommet ( X 16 ). 15: sommet d' une corolle mâle, à 5 divisions régulières ( X 32).16: intérieur d' une corolle mâle, avec ovaire stérile ( ou « faux pédoncule » ) en a, nectaire en n, regme en r, épiregme en e, étamines soudées en manchon en ét. et brosse stigmatique en si ( X 20 ). 17: développement du « manchon des étamines », avec anthéropodes en ap ., caudicules en c et languettes en l ( X 40 ).
Dessins de G. Beauverd.
l' étoile ne sont, dans le cas présent, que des feuilles dites « ramigères », parce qu' à leur aisselle naissent les rameaux courts, parfois subramifiés, qui portent les capitules ou colonies de fleurs.
Ces dernières sont de deux natures ou sexes qui se disposent à l' in du capitule selon un ordre constant: à la périphérie s' épanouissent tout d' abord les fleurs femelles, c'est-à-dire celles qui donnent les semences. Elles sont composées de parties distinctes que nous désignerons comme suit:
a ) l' ovaire, qui deviendra plus tard le fruit dénommé aussi « akène »; sa forme est celle d' un petit fuseau quelque peu tronqué aux deux extrémités et directement inséré sur le « réceptacle » ou plateau du capitule ( fig. I: 10 a, 11 a et 12 a ); b ) l' aigrette, constituée par une couronne de poils disposés à la façon d' un volant pour jeu de raquette: soudés à la base en petit anneau qui persiste jusqu' à la dissémination du fruit, ces poils sont uniformément filiformes de la base au sommet chez les fleurs femelles, ce qui n' est pas le cas pour les fleurs mâles comme nous le verrons plus loin ( fig. I: 7 et 8 ); c ) la corolle, ou petit cylindre blanc verdâtre, excessivement fin, un peu dilaté à la base et terminé au sommet par trois à quatre franges assez irrégulières ( voir fig. I: 9 et 10 ); d ) le pistil, qui, au microscope cela s' entend, présente à la base un nectaire urcéolé, sorte de gobelet minuscule renfermant le nectar recherché par les insectes butineurs, tels que bourdons, abeilles, etc. ( voir fig. I: 11n ); l' intérieur du nectaire abrite un menu filet, nommé « regme », et surmonté d' une protubérance hémisphérique, appelée aussi « épiregme »; c' est de son sommet que s' élève le style généralement plus long que le tube de la corolle et terminé à son extrémité supérieure par les deux branches du stigmate, longues et divergentes ( fig. I: 10 et 11 st ).
Lorsque l' épanouissement des fleurs femelles est entièrement terminé, le centre ou « disque » du capitule se revêt de petites pointes jaunes qui représentent les anthères des fleurs mâles. Ces fleurs mâles sont ainsi désignées, parce qu' elles sont totalement dépourvues de fruits; elles présentent néanmoins les organes suivants:
a ) le pédicelle, qui n' est à proprement parler qu' un ovaire stérile ( absence d' ovule ); il simule un long cône renversé, à section longitudinale fortement concave sur les marges, atténuée à la base et plus élargie au sommet ( fig. I: 16 a ); b ) l' aigrette, qui se distingue de celle des fleurs femelles par ses poils fortement dilatés au sommet, affectant en quelque sorte une forme de petites massues; cet aspect est dû au développement de nombreuses papilles, d' abord courtes et espacées, puis de plus en plus longues et rapprochées selon leur situation vers l' extrémité de l' aigrette; au moment de l' ouverture des anthères, ces papilles se chargent de pollen qu' elles offrent involontairement aux insectes butineurs ( fig. 1:13 et 14 ); c ) la corolle, sensiblement plus grosse que chez les fleurs femelles, et s' en différenciant par sa forme campanulée, évasée au sommet qui est orné de cinq lobes larges et très réguliers ( fig. I: 15 et 16 ); d ) les anthères, au nombre de cinq, insérées au sommet de petits filets qui se libèrent du quart inférieur du tube de la corolle où ils étaient tout d' abord fixés; ces anthères sont alors soudées en manchon par leurs marges, pourvues à la base de deux petits appendices libres, nommés « caudicules », et terminées au sommet par un autre appendice qui a reçu le nom de « languette » à cause de sa forme caractéristique; le pollen est réparti selon deux lignes parallèles partant d' un point situé entre les deux caudicules et dilaté en long fuseau sclérifié, nommé anthéropode; le sommet des deux lignes polliniques aboutit au bas de la languette ( fig. I: 16 ét. et 17 ); e ) le pistil, assez semblable à celui des fleurs femelles, puisqu' il est pourvu à sa base d' un nectaire, d' un regme et d' un épiregme; toutefois, il s' en distingue physiologiquement par son stigmate, dont les deux grosses branches soudées entre elles presque jusqu' au sommet, et pourvues extérieurement de grosses papilles collectrices, sont non seulement inaptes à être fécondées par le pollen, mais fonctionnent au contraire comme appareil collecteur ( nommé « brosse pollinique » ) pour offrir ce pollen aux insectes qui s' en iront le répartir à leur insu sur les stigmates femelles des autres pieds de léontopode du voisinage ( voir fig. I: 16 st. ).
En dehors de ces deux catégories sexuelles de fleurs, réparties dans un même capitule du Leontopodium alpinum, on peut trouver, bien rarement, il est vrai ( en proportion moyenne de 4 à 5‰ selon mes recherches personnelles effectuées durant plusieurs années sur des centaines de pieds provenant des stations les plus diverses de nos Alpes ), des fleurs d' un type différent qui, par leur structure, établissent le passage entre les deux catégories précédentes ou même en exagèrent le caractère d' évolution; ce sont: 1° des fleurs hermaphrodites, à corolles plus grandes et plus régulières que celles des fleurs femelles, à ovaire pourvu d' ovule, mais à an-thères plus ou moins bien formées et garnies de pollen; le stigmate, divisé nettement en deux branches, possède les papilles stigmatiques qui le rendent apte à être fécondé normalement; ces fleurs exceptionnelles, quand elles existent dans un capitule, sont toujours situées au point de transition séparant les rangs extérieurs de fleurs femelles des rangs intérieurs de fleurs mâles; 2 " des ( leurs « subhermaphrodites », à corolles du type franchement femelle, mais agrémentées, à l' intérieur, d' étamines rudimentaires jamais soudées entre elles et d' ailleurs dépourvues de pollen; fertiles, elles ne peuvent être fécondées que par un pollen étranger. Cette catégorie de fleurs, plus rare encore que la précédente, est située dans le dernier rang des fleurs femelles où elle représente un état d' acheminement vers le dimorphisme sexuel plus avancé que chez le type précédent, en ce sens que l' évolution vers le type femelle est presque accomplie par l' ablation partielle des organes mâles; 3° des fleurs neutres 1 ), situées tout au centre du capitule, dans le disque des fleurs mâles: corolles identiques à celles de ces dernières, nectaire relativement développé, stigmate nul et anthères mal formées, généralement destituées de pollen; à cause du nectaire mieux conformé que les autres organes, cette catégorie de fleurs a été considérée comme « appareil d' appel » pour les insectes butineurs; à mon sens, il s' agirait Fig. II. Cycle évolutif théorique de l' anthèse, soit floraison, chez les léontopodes hétérogames.
1: schéma du Leontopodium alpinum avant l' anthèse; aucun capitule n' offre encore de bouton épanoui. 2: id. au début de l' anthèse; le capitule central présente ses premières corolles épanouies, sous forme de fleurs femelles groupées exclusivement à la périphérie. 3: id ., seconde phase de l' anthèse; les fleurs femelles de la périphérie étant toutes fanées, ce sont les fleurs mâles du disque qui s' épanouissent simultanément. 4: id. à la troisième phase de l' anthèse; toutes les fleurs du capitule central étant fanées, les capitules latéraux commencent alors de fleurir simultanément par la périphérie ( stade femelle ). 5: id. à la quatrième phase de l' anthèse; les fleurs périphériques des capitules latéraux étant toutes fanées, ce sont les fleurs mâles du disque qui s' épanouissent ( stade mâle ). 6: id. à la dernière phase; la totalité des fleurs étant fanées, la dissémination des graines va s' opérer, tout d' abord par les rangs périphériques du seul capitule central, ensuite par ceux des capitules latéraux; les fleurs du disque n' ont jamais de graines.
Nota. Ce dispositif exclut, en théorie, la fécondation des fleurs femelles par le pollen d' un même exemplaire donné; en pratique, on peut constater des cas exceptionnels de fécondation des capitules latéraux au stade femelle par le pollen des fleurs du capitule central, à la fin du stade mâle ( observations personnelles ); dans la règle, le Leonlopodium alpinum constitue l' un des exemples les plus typiques d' adaptation exclusive à la fécondation des fleurs par l' intermédiaire des insectes.
Croquis de G. Beauverd.
plutôt de fleurs avortées par l' effet de leur situation à l' extrême région terminale de l' inflorescence ( centre du réceptacle ), le développement normal des fleurs précédemment épanouies ayant épuisé le pouvoir végétatif indispensable à la formation harmonieuse de tous les organes des dernières venues. Ces deux interprétations, d' ailleurs, ne sont pas exclusives et me paraissent pouvoir être admises simultanément par des rapports de cause à effet; au surplus, ces trois formes de « fleurs de transitions » ou de « fleurs avortées » ne sont pas spéciales au genre Leontopodium: elles apparaissent plus fréquemment encore chez quantité d' autres genres de la famille des Composées, en première ligne desquelles il convient de citer les Centaurea et plus encore les Artemisia, qui comprennent les « genépis » de nos montagnards: ces cas d' atavisme sont précieux à noter pour le morphologiste.
Tels sont les principaux détails concernant la structure florale de notre « Edelweiss » alpin; il resterait beaucoup d' autres choses inté- Fig. III. Répartition sexuelle des fleurs chez le genre Leontopodium.
1: inflorescence hétérogame normale, dite « homodynamique » ( tous les capitules d' un même exemplaire considéré présentent une répartition identique de fleurs femelles et de fleurs mâles, par exemple, l' Edelweiss de nos Alpes ). 2: inflorescence hétérogame didynamique, c'est-à-dire acheminée au type dioïque ( le capitule central présente seul une prépondérance marquée de fleurs mâles sur les fleurs femelles rarement absentes; les capitules latéraux offrent une répartition inverse des sexes, avec absence souvent totale de fleurs mâles; exemples; la variété apennine de notre Edelweiss; nombreuses variétés asiatiques; Edelweiss de l' Himalaya, etc. ). 3 et 4: inflorescences dioïques, soit strictement homogames pour tous les capitules d' un même exemplaire donné ( l' exemplaire 3 n' offrira que des capitules à fleurs exclusivement mâles, tandis que les capitules à fleurs exclusivement femelles ne se rencontreront que chez l' exemplaire 4; parfois, une ou deux corolles de sexe adverse apparaîtront dans chacun des capitules d' une même inflorescence; exemples: plusieurs variétés asiatiques de notre Edelweiss européen et nombreuses espèces de Leontopodium asiatiques ).
Croquis de G. Beauverd.
ressantes à signaler sur le rôle des différents organes ci-dessus passés en revue, ainsi que sur une comparaison avec les autres représentants asiatiques du genre Leontopodium, lesquels, pour la plupart, répartissent leur dimorphisme sexuel non plus en deux zones localisées dans un même capitule donné, mais bien uniformément sur deux capitules différents d' un même individu ( par exemple chez le Leonlopodium himalayanum ) ou, plus fréquemment encore, sur tous les capitules de deux exemplaires distincts ( dioicie parfaite des L. leontopodioides, L. subulatum, L. sinense, L. Francheti, etc. ). Mais pour ne pas abuser de l' espace qui nous est concédé, nous ré-sumerons les considérations de cet ordre dans la légende commentant les figures destinées à illustrer le texte, et nous nous bornerons à présenter, dans la planche photographique, quelques-uns des types saillants des autres espèces asiatiques du genre Leontopodium. Pour ceux que d' autres détails essentiels pourraient encore intéresser, nous ne saurions mieux faire que de les renvoyer soit aux articles précédemment publiés par l'«Echo des Alpes » ( année 1881, vol. XVII, p. 286 à 290 ) et l'«Alpina » ( Dr W. Rytz « Die Alpenkönigin », 24. Jahrg. [1916], n° 8, p. 150 à 152 ), soit surtout à la belle notice du D ' ' C. Schröter, publiée dans la 2e édition du « Pflanzenleben der Alpen, eine Schilderung der Hochgebirgsflora » ( Zurich 1924 ), p. 498 à 506, où la bibliographie essentielle du sujet est exposée en détail. Et pour nous résumer, qu' il soit permis d' insister sur le fait qu' après les quatre siècles d' écart qu' embrasse la littérature relative à l' Edelweiss, soit depuis l' époque où cette plante n' était considérée qu' à titre de tisane, jusqu' à notre XXe siècle si fécond en découvertes remarquables dans tous les domaines des recherches humaines, on ne peut certifier qu' une chose à propos d' un simple végétal pourchassé à tel point qu' aucun autre dans les Alpes n' a occasionné autant de morts humaines: c' est qu' il reste encore beaucoup à faire pour pouvoir dire qu' on le connaît à fond.
Car, on ne saurait assez le répéter pour extirper un préjugé tenace: malgré la grande diversité des méthodes appliquées à leurs disciplines respectives, les sciences sont solidaires les unes des autres; ne représentent-elles pas, dans leur ensemble, le plus précieux des apanages humains, celui qui fixe le critère indiscutable de la supériorité de l' homme dans le cadre entier de la création connueAussi bien convient-il, pour quiconque veut mériter sa part de cet apanage, de s' enquérir des diverses phases, soit « stades progressifs » imposés par le temps au programme de la science, et de choisir celui qui convient le mieux à ses moyens.
Dans le plus primitif, le « stade préliminaire » pourrait-on dire, la moindre observation peut fournir sa part de contribution scientifique, depuis celle de l' ouvrier qui participe à la construction et au perfectionnement de l' ou, ou celle du simple indolent qui découvre le principe de la machine à vapeur en se laissant captiver par les trépidations d' une marmite d' eau bouillante, jusqu' à celle du promeneur attentif qui note, au cours d' une ascension, les diverses manifestations extérieures de la vie animale ou végétale.
Un second stade, qu' on pourrait qualifier de « nomenclatural », fait appel à l' esprit de classification pour enrichir un vocabulaire sans l' aide duquel on ne saurait s' entendre sur l' ordre des matières soumises à l' étude; c' est en quelque sorte une phase de « mise en ordre ». Enfin, dans un troisième stade progressif, celui de la « coordination », on consigne les observations du premier stade en utilisant le langage du second, puis on discute les faits, on conclut parfois, sinon l'on édifie un majestueux échafaudage d' hypothèses, plus rarement un monument durable basé sur des lois, en s' aidant pour la circonstance de ces précieux instruments de travail qui font la gloire des laboratoires, des bibliothèques ou des musées modernes.
Sur cette base, il nous sera permis de donner l' éveil sur une erreur communément répandue à l' égard des études concernant le règne végétal, erreur qui se résume dans cette formule qui n' est qu' une boutade commise à l' usage des farceurs: « La botanique est l' art de connaître le plus grand nombre possible de noms de plantes... en langage baroque. » C' est une définition à peine appli- cable à certains « spécialistes » de notre second stade; elle est certainement incompatible avec la réalité qui est à la fois beaucoup plus simple, bien plus sérieuse et infiniment plus complexe: « La botanique est la recherche de la vérité dans tout ce qui a rapport au règne végétal. » Cette constatation est d' importance capitale si l'on songe que c' est à des recherches de cet ordre que l' humanité est redevable de connaître l' une des sources de sa propre existence, cette modeste matière verte qu' est la chlorophylle, laquelle concède au règne végétal le monopole de la fabrication spontanée du charbon et de la régénération de l' oxygène, ceci de concert avec les lois cosmographiques du jour et de la nuit!
Et pour ne s' en tenir qu' aux secrets encore célés dans notre léontopode des Alpes, que ne pourrait-on en obtenir si l'on s' avisait de pousser à fond la solution des problèmes posés par son existenceDepuis ces précurseurs de la médecine qu' étaient les herboristes empiriques des anciens âges, qu' aux biochimistes modernes qui, à l' aide de l' ammoniaque, viennent de nous révéler dans les flavones une importante substance végétale jouant entre autres un rôle défensif contre la radioactivité ultraviolette, ou encore qu' aux géobotanistes qui, dans la discussion des faits relatifs à cette étrange lacune caucaso-ouralienne de l' aire des léontopodes, pourront prêter la main aux géologues pour élucider un point important de l' histoire de notre globe, bien d' autres catégories de chercheurs sont qualifiées pour interroger avec succès notre petite fleur des rochers: blottie dans sa livrée de laine blanche où elle distille un nectar cher à des légions d' insectes, elle brave les vicissitudes du climat meurtrier des hautes régions sans se lasser d' offrir aux investigations humaines un imposant programme de recherches; à lui seul, ce programme mérite de faire pardonner à l' Edelweiss les homicides inconscients dûs à la fascination de son étrange regard.G. Beauverd
Explications de la planche photographique ( v. la planche p. 107 ).
UNE DOUZAINE DE TYPES ASIATIQUES DE LÉONTOPODES.
1: LEONTOPODIUM ALPINUM Cassini ( Europe et Asie; observer la brièveté des rameaux partant du centre de l' inflorescence. 2: L. FRANCHETI Beauverd ( Chine centrale; les feuilles, très étroites, sont glanduleuses et d' un vert olivâtre ). 3: L. DEDE-KENSII Beauverd ( Chine centrale; remarquer les rameaux feuilles vers le milieu de la tige ). 4: L. STRACHEYI C. B. Clarke ( Himalaya, Thibet; petites feuilles obliptiques, ondulées sur les bords ). 5: L. DISCOLOR Beauverd ( Japon; feuilles larges et d' un vert noirâtre dessus, blanches farineuses dessous ). 6: L. LEONTOPODIOIDES ( Willd. ) Beauverd ( Sibérie, Chine septentrionale et orientale; plante dioïque à feuilles radiales éparses, non disposées en étoile; c' est le véritable « Edelweiss de Sibérie » des botanistes, mais non des horticulteurs ). 7: L. HIMALAYANUM DC. ( Himalaya; capitules homo-games: le central à fleurs toutes du même sexe, les latéraux à fleurs toutes de l' autre sexe ). 8: L. SUBULATUM Beauverd ( Chine; plantes dioïques, à très nombreuses petites feuilles étroites et blanches ). 9: L. MONOCEPHALUM Edgeworth var. EVAX Beauverd ( Himalaya; petites feuilles d' un blanc cendré, disposées en rosette; inflores-cence à feuilles bractéales jaune canari ). 10: L. JACOTIANUM Beauverd ( Himalaya, Thibet; pied femelle à inflorescence très laineuse ). 11: id. pied mâle, à inflorescence plus large et brièvement veloutée-cendrée ). 12: L. SINENSE Hemsley ( Thibet oriental et Chine centrale; inflorescence à larges rameaux subramifiés; feuilles d' un jaune verdâtre velouté ).

